embryonic patterning (maternal)
Maternal patterning mutants fall into distinct classes based on the regions of the embryo that are affected (see below).
Some are without an anterior, some are without a posterior, some don't have fronts or backs, and some have no ends. Early studies found multiple maternal genes controlling the development of each of these regions.
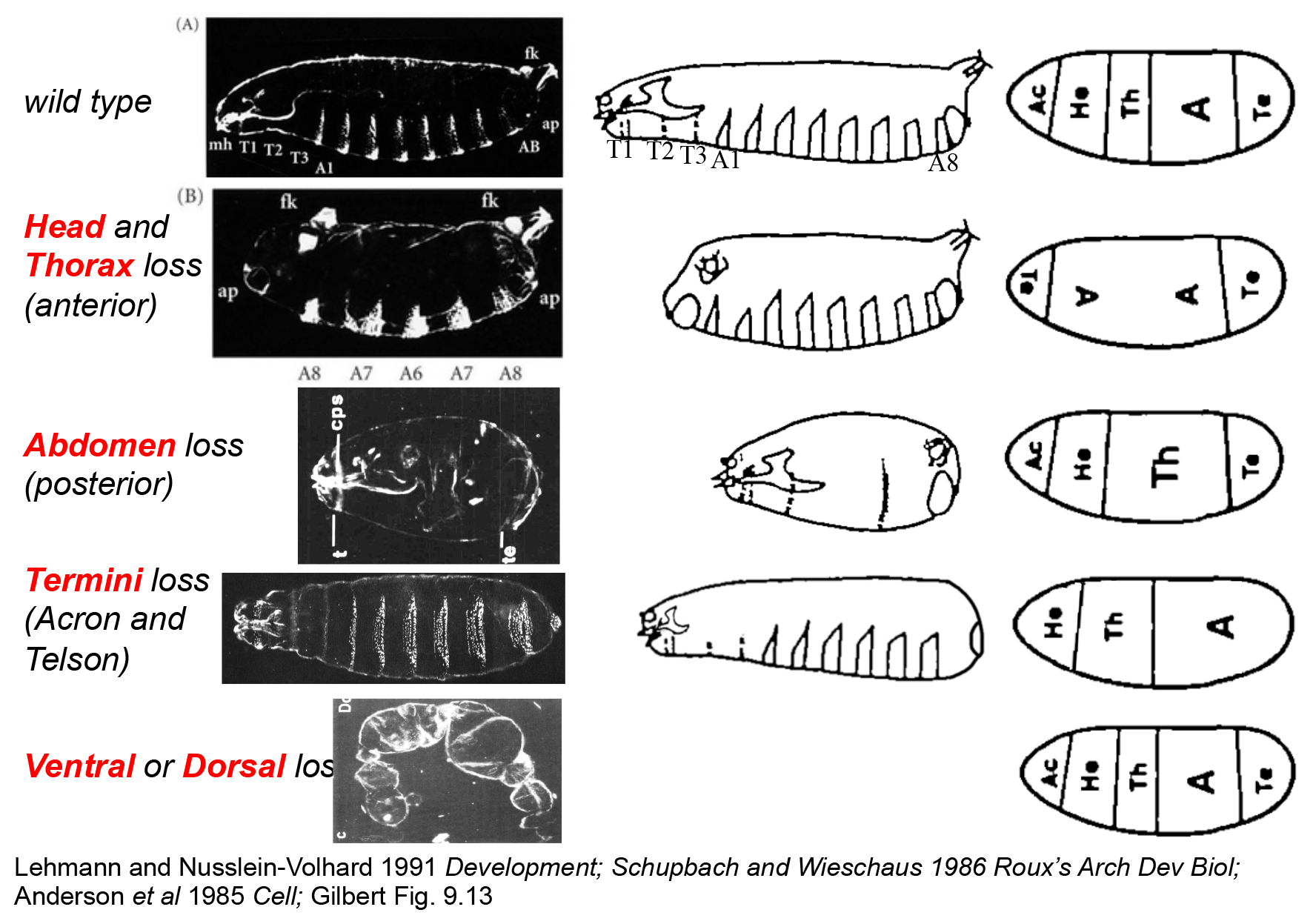
Maternal effect mutant phenotype classes. Actual images of cuticles laid by mutant mothers (left), illustrations of these (middle) and embryonic fate maps interpreted from this work. Acron (A) and Telson (Te) are the termini, Head (He), Thoracic (Th) and Abdomenal (A) regions are shown.
Through a significant body of work performed since these pioneering early studies, we now have a better but still incomplete understanding of these events. A major gap in our knowledge is in how terminal patterning is established.
TERMINAL PATTERNING

A wild type embryo (~2.5h old) showing successful patterning of the termini (red) and dorso-ventral axis (green). These patterning systems rely on restricted cell signalling events and serve as excellent paradigms in which to understand the mechanisms how cell signalling is controlled.
Patterning of the ends of the embryo, or termini, is governed by the localised activation of a receptor called Torso (Tor) that is present on the surface of the embryo. The growth factor known to bind and activate Tor here is called Trunk (Trk) a member of the cysteine-knot superfamily of growth factors. Upon binding of Trk to Tor, a signal is transduced from the via the highly conserved Ras / mitogen activated protein kinase phosphorylation cascade, that eventually leads to the gene activation in the zygote and proteins that determine the fate of the termini are made.

Eggs laid by terminal class mutant mothers all fail to develop structures posterior to the A7 segment, and have a disrupted anterior.
the terminal class phenotype
One of the interesting classes of mutants found from these screens was one where loss of the gene's function in the mother lead to embryos that develop with disrupted heads and no tails. This is distinct from disruption of the maternal other patterning systems.

Activation of the Torso cell signalling pathway at each end of the embryo drives the patterning program in these regions.
the molecules involved
Cloning and sequencing of the terminal class genes in the late 1980s and early 1990s revealed that terminal pattern formation was controlled by a Receptor Tyrosine Kinase cell signalling pathway.
- torso (tor) encodes the receptor.
- trunk (trk) encodes the growth factor that binds to and activates Torso.
- Several eggshell proteins are required: female sterile (fs)Nasrat (N), fs(1)polehole (ph) and closca (clos). These are large structural components of the innermost layer of the fly eggshell - the vitelline membrane.
- The highly conserved Ras / MAPK phosophorylation cascade transmits the signal from Receptor to the nucleus to control the expression of Zygotic genes (tailless and huckebein).
- Torso-like (tsl) encodes a relative of the Perforin-like protein superfamily known to form large oligomeric pores on target cell membranes. The function of Torso-like here remains a mystery.
Torso-like holds the key
Torso-like is unique amongst the terminal class genes in that it is the only known terminal class gene with an expression pattern that is restricted to the ends of the embryo. All other factors are thought to be present throughout the embryo. Forced expression of Torso-like all around the embryo causes a great expansion of the terminal region at the expense of central segments, known as spliced. These studies suggest that Torso-like determines where terminal patterning occurs.
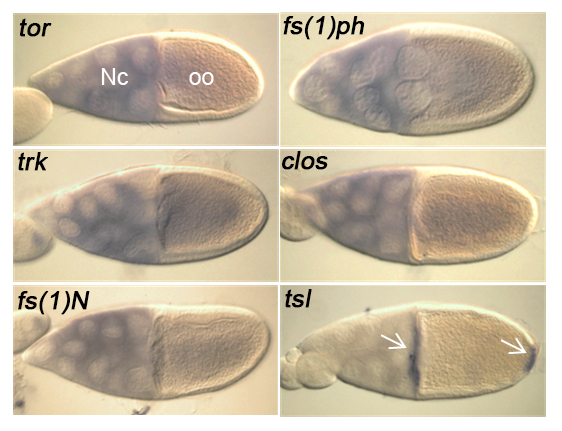
Expression of the terminal class genes in the ovary (egg chamber). The nurse cells (Nc) load the oocyte (oo) with gene products for embryonic development. Note that tsl expression is limited to cells at each end of the oocyte.

Embryo laid by female expressing tsl in all ovarian follicle cells causes loss of central segments and enlarged termini.